
Two Endoceras giganteum forage on the seafloor, while a hapless Isotelus gigas gets a little too close. Art by Prehistorica, reproduced here with permission.
The orthoceratoid Endoceras is the both the longest extinct cephalopod and largest Paleozoic invertebrate. A shell of E. giganteum (MCZ unnumbered) from the Late Ordovician of New York is the current record-holding specimen. As preserved it measures 3 meters in length, with an estimated complete length of 5.73 meters (Teichert & Kummel, 1960; Klug et al. 2015). There is even an anecdotal report of a 9.14 meter shell that was destroyed in the field (Flower, 1955a). Despite its impressive size, Endoceras is not particularly well-represented in paleoart. The few depictions are plagued by the lack of research found in most art of extinct cephalopods.
By incorporating data from both orthoceratoids and extant nautiluses, I have attempted a more rigorous restoration of this taxon. This will be the first in a series of posts about the life appearances of fossil cephalopods, intended as reference material for paleoartists. I am following the classification scheme for nautiluses and relatives proposed by King & Evans (2019), albeit retaining the traditional spellings of clade names to avoid confusion. I am also using the dichotomy of Palcephalopoda (all cephalopods closer to living nautiluses) and Neocephalopoda (all cephalopods closer to living coleoids) proposed by Lehmann & Hillmer (1980/1983).
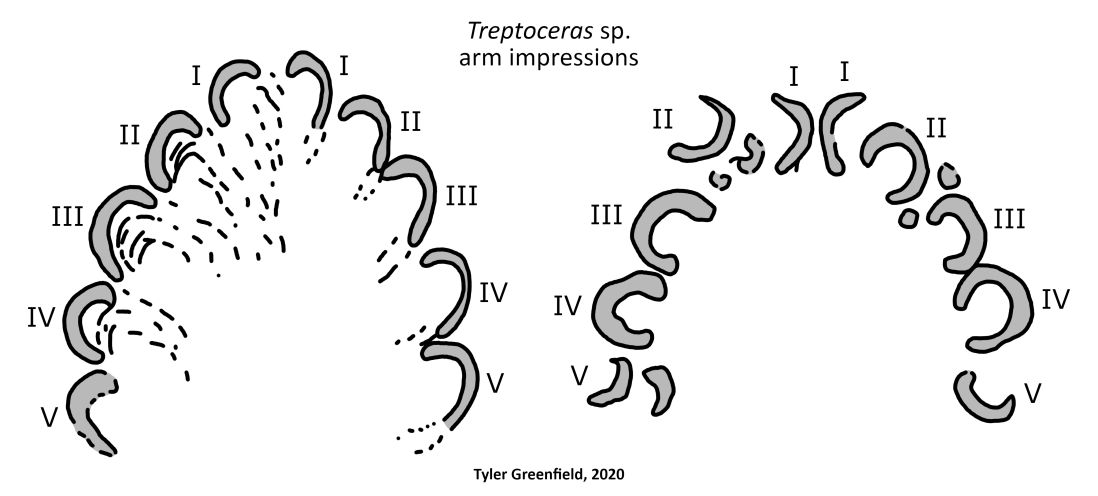
Impressions from Treptoceras labelled with corresponding arm pairs, redrawn from Flower (1955b).
While no soft tissues from Endoceras are known, some exceptional fossils from relatives give insight into how it would have looked. In particular, two trace fossils from the Late Ordovician of Ohio assigned to the orthocerid Treptoceras can be used to reconstruct the arm crown. These impressions, which represent the animal resting or feeding on the seafloor, indicate 10 arms of roughly equal length. Although only traces, they can be confidently referred to an orthocerid since no other contemporary organism matches their morphology and they occur in the same strata as shell trails (Flower, 1955b). They conform with the 10-armed ancestral condition hypothesized for cephalopods and with the development of nautiluses. Adult nautiluses have around 90 small arms, but they all originate from 10 arm buds in the embryonic stage (Shigeno et al. 2008).
Mehl (1984) also reported a specimen of the orthocerid Michelinoceras from the Late Silurian of Bolivia with 2 arm imprints, which he identified as tentacles. However, this is dubious because the other arms are not preserved for comparison. Tentacles are only found in extant decabrachians and the other, more complete orthocerid impressions lack them. Additionally, I am uncertain if they are actually the remains of arms or instead a taphonomic artifact. Thus it remains most likely that orthoceratoids including Endoceras had 10 arms without a pair differentiated into tentacles.
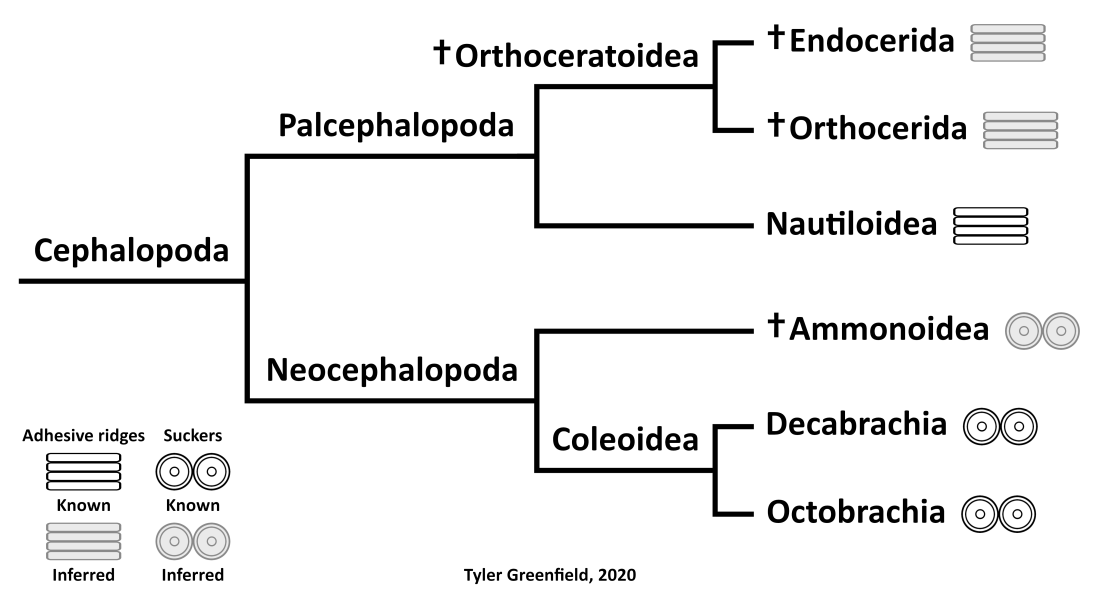
Phylogenetic distribution of adhesive structures in the Cephalopoda.
The adhesive structures on the undersides of the arms are a matter of speculation. The recent discovery of decabrachian-like arm hooks in ammonoids (Kruta et al. 2020) means they may have had suckers like coleoids, which in turn would imply that suckers evolved in the Neocephalopoda. Nautiluses on the other hand have ridges on their arms that secrete a strong glue (von Byern et al. 2012). Using phylogenetic bracketing suggests other palcephalopods including orthoceratoids had adhesive ridges too. Presumably the ridges would have been larger on the more well-developed arms of orthoceratoids to aid in prey capture.
The available data on the arms of orthoceratoids contradicts the filter-feeding ecology hypothesized for endocerids by Mironenko (2020). The only confirmed filter-feeding cephalopod, the vampire squid Vampyroteuthis infernalis, uses velar filaments (a modified arm pair) in combination with suckers and cirri for feeding (Hoving & Robison, 2012). As evidenced by the trace fossils, orthoceratoids did not have an arm pair modified into tentacles or filaments. Suckers are only known in coleoids and cirri in octobrachians, so it is unlikely that orthoceratoids would have possessed either. Predating and scavenging on benthic prey is a more plausible ecological niche for endocerids.
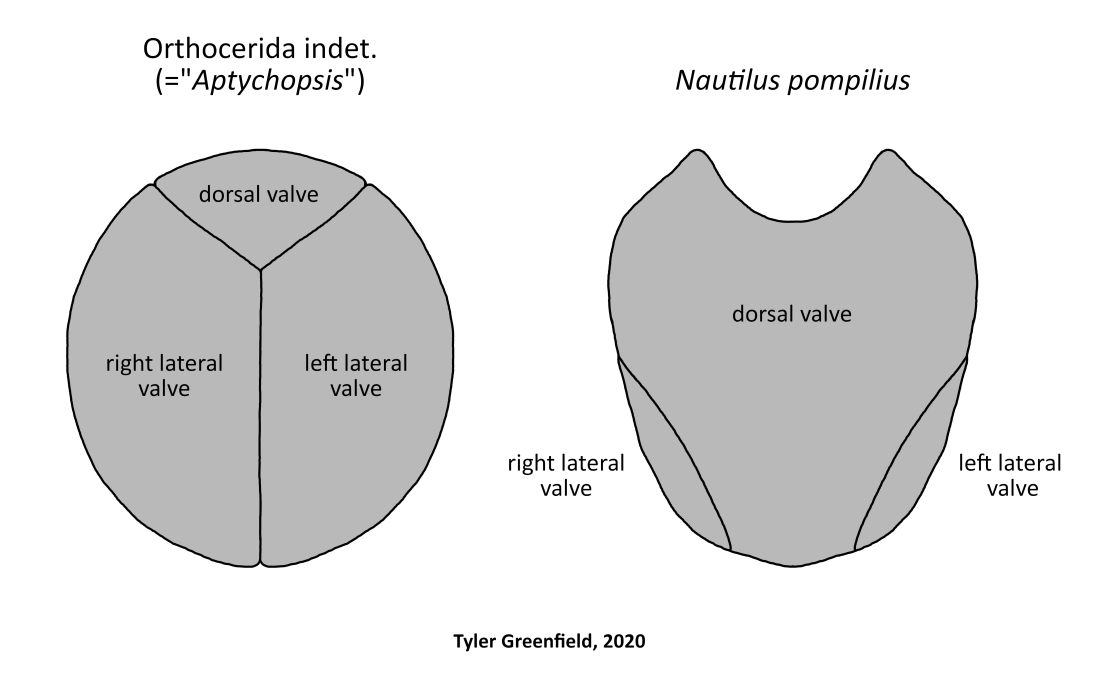
Comparison between the opercula of orthocerids and modern nautiluses showing homologous elements. “Aptychopsis” redrawn from Dzik (1981) and Nautilus redrawn from Dean (1901); terminology from Turek (1978).
There is evidence that all orthoceratoids had an operculum on top of the head as in nautiluses. Unlike the hood-shaped operculum of nautiluses, opercula found with orthocerid shells (referred to the parataxon “Aptychopsis“) are relatively flat and oval-shaped. This morphological difference is to accommodate the shape of the shell opening, as the purpose of the operculum is to cover the opening for protection. The homology between nautilus and orthocerid opercula is demonstrated in that they are both composed of three interlocking valves (Turek, 1978; Holland et al. 1978; Stridsberg, 1984). “Aptychopsis” valves have alternately been interpreted as jaws by some workers (e.g. Dzik, 1981), but this is doubtful considering they do not resemble any other cephalopod jaw types.
Like the adhesive structures, reconstructing the eyes of orthoceratoids must rely on phylogenetic bracketing. Nautiluses have a simple pinhole-type eye without a lens, which is a primitive characteristic and may represent the ancestral cephalopod condition (Sasaki et al. 2010). Camera-type eyes with lenses are known in coleoids, and the presence of coleoid-like eye capsules in ammonoids indicates they had them as well (Klug et al. 2012). It seems that, like suckers, camera-type eyes evolved in the Neocephalopoda. Bracketing would suggest that orthoceratoids and other palcephalopods instead had pinhole-type eyes. If this is the case, it means that Endoceras may have had poor vision like nautiluses and relied on other senses for finding prey.
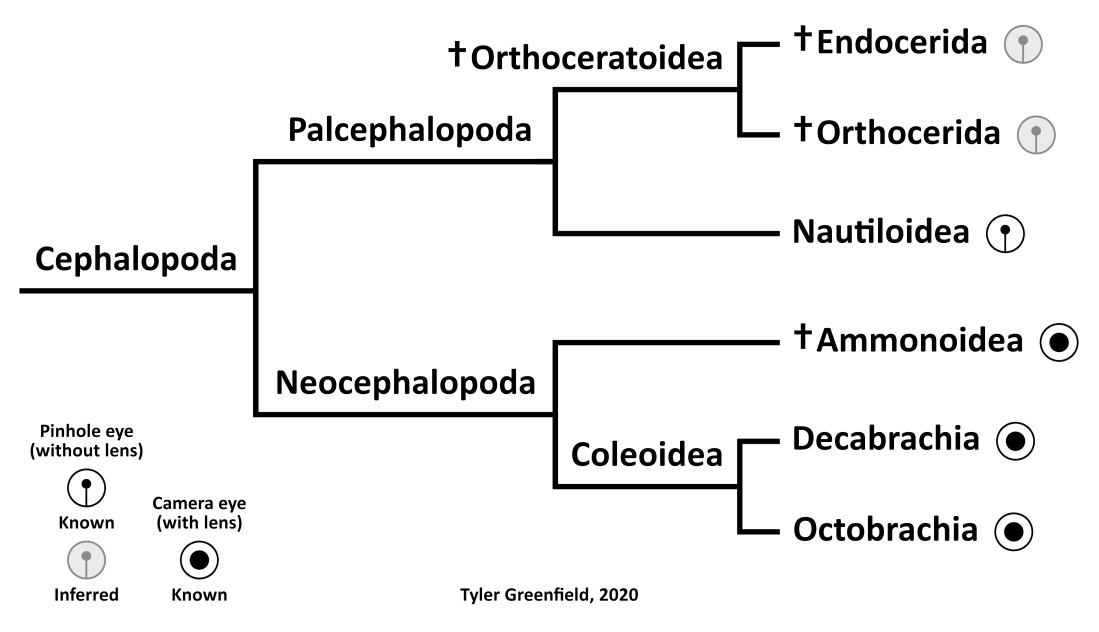
Phylogenetic distribution of eye types in the Cephalopoda.
Preserved color markings are known from a large number of palcephalopods (see Kobluk & Mapes, 1989 for a partial list); note that these do not show the original colors, only the patterns. Markings on a shell of Endoceras proteiforme are described as “cancellated” by Marsh (1869), but unfortunately this specimen is not illustrated and is currently lost. Even though this report is anecdotal, another endocerid Anthoceras vaginatum has similar mottled markings (Kröger, 2012) so this coloration is still plausible. Exactly what function it would have served is unknown, with camouflage/mimicry or display being most likely. On a side note, the color of the trilobite Isotelus in the featured artwork is also based on preserved patterns (Wells, 1942).
A recent study using 3D modeling determined that orthocerids and endocerids maintained a vertical shell orientation while suspended in the water column (Peterman et al. 2019). This is more evidence against filter feeding, as horizontal movement conducive to filtering would have been restricted. Movement on the seafloor was more varied, with the aforementioned arm impressions showing the shell held vertically, while other trace fossils show movement of the shell horizontally through the sediment (Flower, 1955b). Taken together, the shell orientation and preserved interactions with the seafloor indicate that orthoceratoids were generally benthic feeders.
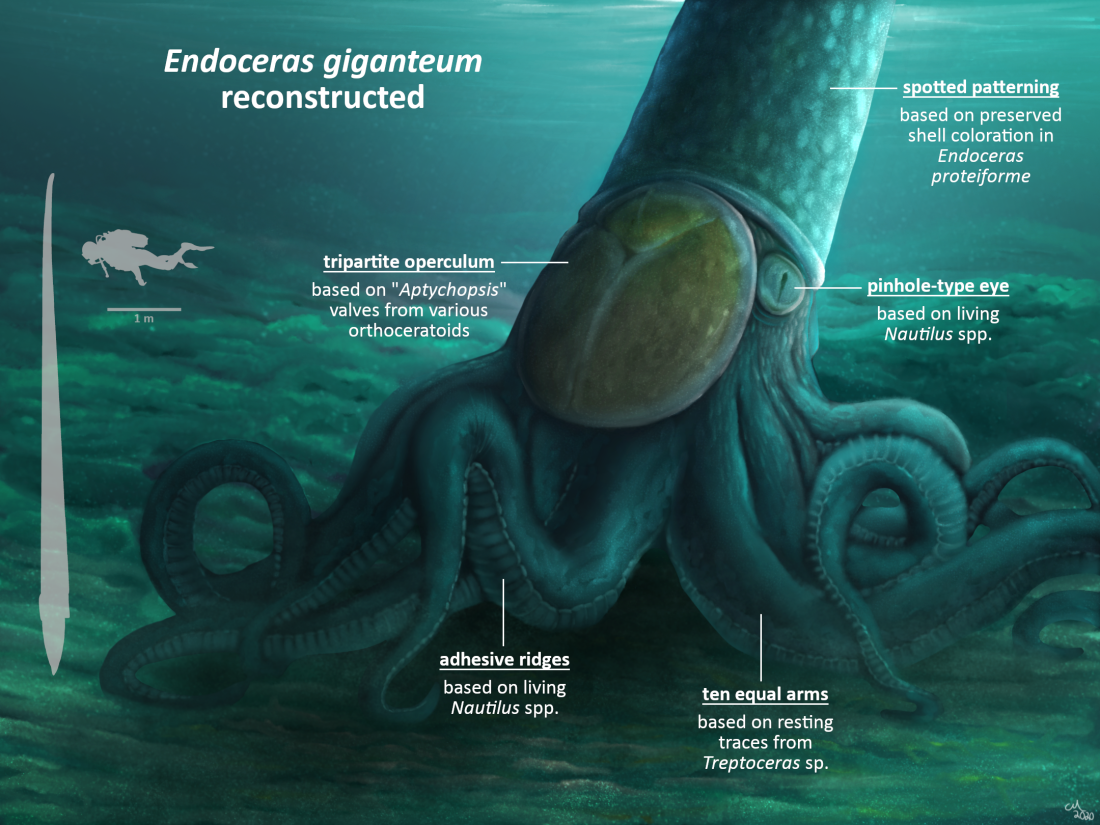
A summary of the information used to reconstruct Endoceras in this post, with art again by Prehistorica.
All of this information was utilized by paleoartist Prehistorica to produce the life reconstruction of Endoceras for this blog. This is probably the most accurate depiction to date, going against traditional ones that are either too nautilus-like or too coleoid-like. Of course some of the features are conjectural, but they are based on both fossil evidence and phylogenetic bracketing, and not pure artistic license. Hopefully this post (and upcoming ones in this series) will encourage other artists to research extinct cephalopods and illustrate them more accurately.
Special thanks go to Prehistorica who created the awesome artwork for this blog. Go check out his Twitter where he regularly posts his paleoart, especially of Cambrian lobopods.
Addendum (7/24/2020)

A new Fascifodina from the Georgian Bay Formation of Toronto. Photo by Joe Moysiuk, used with permission.
The possible orthoceratoid arm traces described by Flower (1955) were later given their own ichnogenus/species, Fascifodina floweri, by Osgood (1970). Osgood reinterpreted them as being burrows, but I am skeptical of this considering how well they match the hypothesized arm crown of orthoceratoids. Joe Moysiuk recently discovered an additional Fascifodina from the Georgian Bay Formation of Toronto, Canada. Interestingly this formation is roughly coeval with Corryville Formation in Ohio where the original Fascifodina are from, and the orthoceratoid fauna is similarly dominated by Treptoceras. Exactly what implications this has is yet unknown, but new fossils like this could settle the true affinity of Fascifodina.
Addendum (7/29/2021)

A Pseudogygites with a bite mark and the hypothetical endocerid beak that made it. The Pseudogygites is redrawn and modified from the reconstruction in Ludvigsen (1979), with the antennae redrawn and modified from the reconstruction of Isotelus in Raymond (1920). The endocerid beak is redrawn and modified from a Nautilus beak in Kanie (1982).
It turns out that there is direct evidence for endocerid predation on trilobites, which further supports that they were benthic feeders. A bite mark was found on a Pseudogygites latimarginatus, a close relative of Isotelus, from the Late Ordovician Lindsay Formation of Ontario. The bite is triangular and best matches the shape of a cephalopod beak; it did not break the exoskeleton, but instead deformed the pleural lobes and pygidium. This probably occurred because the exoskeleton was still soft and pliable soon after moulting. Although the trilobite initially survived the attack, it died from the injury before it could moult again (Rudkin, 1985). The large endocerids Endoceras proteiforme and Cameroceras trentonense are present in the Lindsay Formation (Liberty, 1969) and are the most likely culprits.
Addendum (4/14/2022)

A phylogenetic tree of basal Cephalopoda based on the analysis of Pohle et al. (2022).
A new phylogenetic analysis of early Paleozoic cephalopods by Pohle et al. (2022) has found Palcephalopoda to be paraphyletic and Orthoceratoidea (as traditionally conceived including Endocerida) to be polyphyletic. Endoceratoidea is the sister group to Multiceratoidea, which may include extant nautiluses (in which case it would be renamed Nautiloidea). Endoceratoidea+Multiceratoidea is in turn the sister group to Orthoceratoidea, which includes Neocephalopoda. As a result, the classification scheme used in this post is now outdated. However, the conclusions about endocerid soft tissues still stand given they are still related to nautiluses and orthocerids.
Addendum (6/18/2023)
Destin Bogart of E.D.G.E. has made a good video based on this post.
Next Entry: Enchoteuthis
References
- Dean, B. (1901). Notes on living Nautilus. The American Naturalist, 35(418), 819-837.
- Dzik, J. (1981). Origin of the Cephalopoda. Acta Palaeontologica Polonica, 26(2), 161-191.
- Flower, R.H. (1955a). Status of endoceroid classification. Journal of Paleontology, 29(3), 329-371.
- Flower, R.H. (1955b). Trails and tentacular impressions of orthoconic cephalopods. Journal of Paleontology, 29(5), 857-867.
- Holland, B., Stridsberg, S., & Bergström, J. (1978). Confirmation of the reconstruction of Aptychopsis. Lethaia, 11(2), 144.
- Hoving, H.J.T., & Robison, B.H. (2012). Vampire squid: detrivores in the oxygen minimum zone. Proceedings of the Royal Society B, 279(1747), 4559-4567.
- Kanie, Y. (1982). Cretaceous tetragonitid ammonite jaws: a comparison with modern Nautilus jaws. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 125, 239-258.
- King, A.H., & Evans, D.H. (2019). High-level classification of the nautiloid cephalopods: a proposal for the revision of the Treatise Part K. Swiss Journal of Palaeontology, 138(1), 65-85.
- Klug, C., De Baets, K., Kröger, B., Bell, M.A., Korn, D., & Payne, J.L. (2015). Normal giants? Temporal and latitudinal shifts of Palaeozoic marine invertebrate gigantism and global change. Lethaia, 48(2), 267-288.
- Klug, C., Riegraf, W., & Lehmann, J. (2012). Soft-part preservation in heteromorph ammonites from the Cenomanian-Turonian boundary event (OAE 2) in north-west Germany. Palaeontology, 55(6), 1307-1331.
- Kobluk, D.R., & Mapes, R.H. (1989). The fossil record, function, and possible origins of shell color patterns in Paleozoic marine invertebrates. PALAIOS, 4(1), 63-85.
- Kröger, B. (2012). The “Vaginaten”: the dominant cephalopods of the Baltoscandian Mid Ordovician endocerid limestone. GFF, 134(2), 115-132.
- Kruta, I., Bardin, J., Smith, C.P.A., Tafforeau, P., & Landman, N.H. (2020). Enigmatic hook-like structures in Cretaceous ammonites (Scaphitidae). Palaeontology, 63(2), 301-312.
- Lehman, U., & Hillmer, G. (1983). Fossil Invertebrates (J. Lettau, Trans.). Cambridge University Press. (Original work published 1980)
- Liberty, B.A. (1969). Palaeozoic geology of the Lake Simcoe area, Ontario. Geological Survey of Canada Memoir, 355, 1-201.
- Ludvigsen, R. (1979). The Ordovician trilobite Pseudogygites Kobayashi in eastern and Arctic North America. Life Sciences Contributions, Royal Ontario Museum, 120, 1-41.
- Marsh, O.C. (1869). On the preservation of color in fossils from Paleozoic formations. Proceedings of the American Association for the Advancement of Science, 17, 325-326.
- Mehl, J. (1984). Radula und fangarme bei Michelinoceras sp. aus dem Silur von Bolivien [Radula and arms of Michelinoceras from the Silurian of Bolivia]. Paläontologische Zeitschrift, 58(3-4), 211-229.
- Mironenko, A.A. (2020). Endocerids: suspension feeding nautiloids? Historical Biology, 32(2), 281-289.
- Osgood, R.G. Jr. (1970). Trace fossils of the Cincinnati area. Palaeontographica Americana, 6(41), 281-444.
- Peterman, D.J., Barton, C.C., & Yacobucci, M.M. (2019). The hydrostatics of Paleozoic ectocochleate cephalopods (Nautiloidea and Endoceratoidea) with implications for modes of life and early colonization of the pelagic zone. Palaeontologia Electronica, 22(2), Article 24A. https://doi.org/10.26879/884
- Pohle, A., Kröger, B., Warnock, R.C.M., King, A.H., Evans, D.H., Aubrechtová, M., Cichowolski, M., Fang, X., & Klug, C. (2022). Early cephalopod evolution clarified through Bayesian phylogenetic inference. BMC Biology, 20, Article 88. https://doi.org/10.1186/s12915-022-01284-5
- Raymond, P.E. (1920). The appendages, anatomy, and relationships of trilobites. Memoirs of the Connecticut Academy of Arts and Sciences, 7, 1-169.
- Rudkin, D. (1985). Exoskeletal abnormalities in four trilobites. Canadian Journal of Earth Sciences, 22(3), 479-483.
- Sasaki, T., Shigeno, S., & Tanabe, K. (2010). Anatomy of living Nautilus: reevaluation of primitiveness and comparison with Coleoidea. In K. Tanabe, Y. Shigeta, T. Sasaki, & H. Hirano (Eds.), Cephalopods: Present and Past (pp. 35-66). Tokai University Press.
- Shigeno, S., Sasaki, T., Moritaki, T., Kasugai, T., Vecchione, M., & Agata, K. (2008). Evolution of the cephalopod head complex by assembly of multiple molluscan body parts: evidence from Nautilus embryonic development. Journal of Morphology, 269(1), 1-17.
- Stridsberg, S. (1984). Aptychopsid plates – jaw elements or protective operculum. Lethaia, 17(1), 93-98.
- Teichert, C., & Kummel, B. (1960). Size of endoceroid cephalopods. Breviora, 128, 1-7.
- Turek, V. (1978). Biological and stratigraphical significance of the Silurian nautiloid Aptychopsis. Lethaia, 11(2), 127-138.
- von Byern, J., Wani, R., Schwaha, T. Grunwald, I., & Cyran, N. (2012). Old and sticky – adhesive mechanisms in the living fossil Nautilus pompilius (Mollusca, Cephalopoda). Zoology, 115(1), 1-11.
- Wells, J.W. (1942). Supposed color-markings in Ordovician trilobites from Ohio. American Journal of Science, Series 5, 240(10), 710-713.

You must be logged in to post a comment.